
Abstract
Mutations in PINK1 and Parkin are two of the main causes of recessive early-onset Parkinson’s disease (PD). We generated human induced pluripotent stem cells (hiPSCs) from fibroblasts of a 64-year-old male patient with a homozygous ILE368ASN mutation in PINK1, who experienced disease onset at 33 years, and from fibroblasts of a 61-year-old female patient heterozygous for the R275W mutation in Parkin, who experienced disease onset at 44 years. Array comparative genomic hybridization (aCGH) determined genotypic variation in each line. The cell lines were successfully used to generate midbrain dopaminergic neurons, the neuron type primarily affected in PD.
1. Resource table
| Unique stem cell lines identifier | LCSBi002-B LCSBi002-C LCSBi004-A LCSBi004-B |
| Alternative names of stem cell lines | ND40066-PINK1-ILE368ASN-clone 7 (LCSBi002-B)ND40066-PINK1-ILE368ASN-clone 8 (LCSBi002-C)ND29369-PARKIN/PARK2-R275W-clone 1 (LCSBi004-A) ND29369-PARKIN/PARK2-R275W-clone 4 (LCSBi004-B) |
| Institution | Gladstone Institutes, CA, USA |
| Contact information of distributor | Gabriela Novak gabriela.novak@alumni.utoronto.ca Alexander Skupin alexander.skupin@uni.lu |
| Type of cell lines | iPSC |
| Origin | Human |
| Cell Source | Fibroblasts |
| Clonality | Clonal |
| Method of reprogramming | CytoTune-iPS Sendai Reprogramming kit (ThermoFisher Scientific). transgenes/vectors used:CytoTuneTM 2.0 KOS (Sox2) CytoTuneTM 2.0 hc-Myc CytoTuneTM 2.0 hKlf4 Clearance was confirmed using Scorecard, which detects the presence of Sendai virus, passage number is listed in Fig. 1d. |
| Multiline rationale | Mutations in two genes known to interact, leading to the same disease |
| Gene modification | YES |
| Type of modification | Familial, spontaneous mutation |
| Associated disease | Parkinson’s disease |
| Gene/locus | PINK1 (PARK6) ILE368ASN 1p36.12 PARKIN/PARK2-R275W 6q26 |
| Method of modification | N/A |
| Name of transgene or resistance | N/A |
| Inducible/constitutive system | N/A |
| Date archived/stock date | 2016 |
| Cell line repository/bank | https://hpscreg.eu/cell-line/LCSBi002-B https://hpscreg.eu/cell-line/LCSBi002-C https://hpscreg.eu/cell-line/LCSBi004-A https://hpscreg.eu/cell-line/LCSBi004-B |
| Ethical approval | Samples were collected in accordance with the US Government guidelines and are subject to MTA issued by Coriell Institute for Medical Research NINDS Cell Repository. The iPSC reprogramming protocol was approved by the Committee on Human Research at the University of California, San Francisco. |
2. Resource utility
Parkinson’s disease (PD) leads to the death of midbrain dopaminergic (mDA) neurons. This poses a major obstacle to the study of the disease. Differentiating neurons from iPSCs of patients who carry PD-related mutations provides an almost unlimited source of mDA neurons and an invaluable resource for the study of PD.
3. Resource details
Parkinson’s disease (PD) is one of the most prevalent neurodegenerative disorder, second only to Alzheimer’s disease. It is characterized by the loss of midbrain dopaminergic (mDA) neurons (Ando et al., 2017, Madsen et al., 2021, Novak et al., 2022, Zhu et al., 2018). The pathological mechanisms of PD are only partly understood, and there is no treatment able to reverse its progression. By the end stage of the disease, about 90% of mDA neurons die, posing a major obstacle to the study of human mDA neurons affected by PD. Technological advancements in somatic cell reprogramming into iPSCs and directed differentiation into mDA neurons has created an essential resource for PD research. We can now generate patient-based iPSCs from skin cells of PD patients who carry PD-associated mutations (Novak et al., 2022). Hence, the mechanism of disease development due to individual mutations can be studied in genetic backgrounds shown to be permissive to disease development. The aCGH analysis was used to identify the gene variants each patient carries, to allow for the investigation of the effects of genetic background (Supplement and a resource details file “CGH data”) (see Table 1 and 2).
Table 1. Summary of lines.
| iPSC line names | Abbreviation in figures | Gender | Age | Ethnicity | Genotype of locus | Disease |
|---|---|---|---|---|---|---|
| ND40066-PINK1-clone 7 (LCSBi002-B) | ND40066-7 | Male | 64 | Caucasian | ILE368ASN | Parkinson’s disease |
| ND40066-PINK1-clone 8 (LCSBi002-C) | ND40066-8 | |||||
| ND29369-PARKIN-clone 1 (LCSBi004-A) | ND29369-1 | Female | 61 | Hispanic | R275W | Parkinson’s disease |
| ND29369-PARKIN-clone 4 (LCSBi004-B) | ND29369-4 |
Table 2. Characterization and validation.
| Classification | Test | Result | Data |
|---|---|---|---|
| Morphology | Photography | normal | Fig. 1 panel a and Supplement |
| Phenotype | Qualitative analysis: Immunocytochemistry | Confirmed by staining for pluripotency markers: Oct4 & Tra-1-60 | Fig. 1 panel a Supplement |
| Quantitative analysis: RT-qPCR | Determined by expression of iPSC-specific transcripts via qPCR (OCT4, NANOG, SOX2), by Scorecard and by FACS (staining for SSEA4 with more than 90% cells positive). | Fig. 1 panel b Fig. 1 panel d FACS – Supplement | |
| Genotype | Karyotype | ND 40066: 46,XY normal human male karyotype ND27760: 46, XX, normal female karyotype, also confirmed in iPSCs. | Attached as supplementary figure |
| Identity | Array comparative genomic hybridization STR analysis | aCGH Probes: Pass SNP Probes: Pass | Attached as supplementary figure |
| submitted | Submitted in archive with journal * | ||
| Mutation analysis | Sequencing | Homozygous PINK1 mut. Heterozygous PARKIN mut. | Fig. 1 panel c and Supplement |
| Microbiology and virology | Mycoplasma | Mycoplasma testing: Negative | Supplement |
| Differentiation potential | Scorecard | Embryonic bodies show ability to differentiate into all three lineages | Fig. 1 panel d, and Supplement |
| Donor screening (OPTIONAL) | HIV 1 + 2 Hepatitis B, Hepatitis C | All samples negative for HIV 1, Hepatitis B, & Hepatitis C | Supplement |
| Genotype additional info (OPTIONAL) | Blood group genotyping | N/A | |
| HLA tissue typing | N/A |
*Parental line was not included, however, a full aCGH analysis and a STR analysis were performed for all ND40066 clones and their profiles match. and we confirmed the presence of the mutation by sequencing all ND40066 clones. Both ND29369 clones were confirmed to carry the PARKIN R275W mutation and their STR profiles match.
When mitochondria become damaged, PINK1 recruits Parkin to the outer mitochondrial membrane, which initiates mitophagy (Ando et al., 2017). The fact that a mutation in either PINK1 or Parkin leads to PD underscores the importance of this pathway in PD development. The PINK1 ILE368ASN mutation interferes with mitophagy by reducing the interaction of PINK1 with its chaperone, HSP90, which destabilizes PINK1 at the mitochondrial membrane, and by reducing its ubiquitin kinase activity through deformation of its substrate binding pocket (Ando et al., 2017). Mutations in Parkin are one of the most common causes of recessive juvenile onset Parkinson’s disease and of sporadic early-onset PD (Zhu et al., 2018). However, Parkin participates in multiple pathways within the cell, making it challenging to identify the specific effects of Parkin mutations in PD (Zhu et al., 2018).
Fibroblasts homozygous for the PINK1 mutation ILE368ASN were obtained from a 64-year-old male patient with disease onset at 33 years (ND40066, Coriell Institute), and fibroblasts heterozygous for the R275W missense mutation in exon 7 of Parkin, which falls within the RING finger 1 domain of the Parkin protein (Madsen et al., 2021), were obtained from a 61-year-old female patient with disease onset at 44 years (LCSBi004, ND29369) (Madsen et al., 2021). Fibroblasts were reprogrammed using the Sendai virus reprogramming method, which should not introduce changes into the genome. Expression of the iPSC markers Oct3/4 and Tra-1-60 was shown by immunocytochemistry, alongside transmitted light (TL) images of iPSC colonies (Fig. 1a, Supplement) and the expression of iPSC status markers OCT4 (POU5F1), SOX2 and NANOG was determined by qPCR (Fig. 1b). In Fig. 1b, the 29542-3 and 29542-7 stand for the cell line ND29542, clones 3 and 7, respectively, which are previously published well classified clones of the ND29542 cell line (Novak et al., 2021). The 40066-7 and 40066-8 stand for the PINK1 mutation-carrying cell line ND40066 clones 7 and 8, respectively. The 29369-1 and 29369-4 stand for the PARKIN mutation-carrying cell line ND29369 clones 1 and 4, respectively. Diff 1, 2 and 3 are samples of neurons differentiated from ND40066-8 via protocol published in (Novak et al., 2022), to illustrate loss of OCT4 (POU5F1), SOX2 and NANOG upon differentiation. The ND40066 clone 8 is in itself a well-classified cell line (Novak et al., 2022). The iPSC status of this cell line was determined at the single cell level, as described in (Novak et al., 2022). Furthermore, the iPSC status of the ND40066-7, ND40066-8, ND29369-1 and ND29369-4 cell lines was determined by comparison of the expression of an iPSC marker panel by these cells to a library of well classified iPSC cell lines using Scorecard analysis (Supplement, section “Scorecard – iPSC marker expression”). Presence of the mutations was confirmed by sequencing (Fig. 1c, Supplement). The iPSC status and trilineage differentiation capacity of the clones was determined using Scorecard (Fig. 1d, Supplement). Scorecard uses TaqMan probes to several iPSC status markers to determine the iPSC status of each cell line. To analyse each cell line’s trilineage potential, the expression of markers for each of the three early germ layers, the endoderm, the ectoderm, and the mesoderm, is determined. These layers are generated by allowing the iPSCs to spontaneously differentiate into embryonic bodies (EBs). Genotypic variation was determined by aCGH analysis (Supplement). It should be noted that comparative genomic hybridization (CGH) array does not detect translocations or inversions, alterations in chromosome structure, mosaicism or polyploidy. Both cell lines were screened and found negative for HIV 1, Hepatitis B, and Hepatitis C and mycoplasma (Supplement).
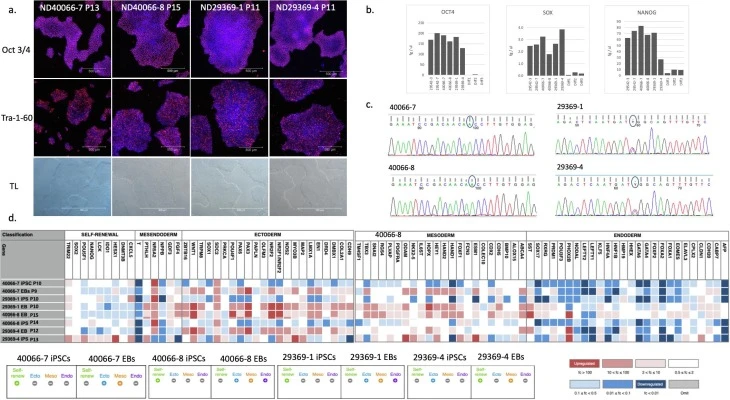
Fig. 1.
Both cell lines were used to successfully generate mDA neurons in our laboratory, which were analysed by single cell RNA seq analysis (Novak et al., 2022).
4. Materials and methods
Fibroblasts were obtained from the Coriell Institute and were cultured as described in recently published methods section pertaining to cells processed in parallel (Novak et al., 2021).
4.1. Karyotype and aCGH analysis
Live adherent fibroblasts of the ND40066 cell line were karyotyped and the genotypic variation of the iPSCc clones 7 and 8 of this cell line was analysed using Array Comparative Genomic Hybridization (aCGH), a high resolution karyotype analysis for the detection of unbalanced structural and numerical chromosomal alterations (Supplement) (Cell Line Genetics, Madison, WI, USA) as described previously (Novak et al., 2021). Live adherent fibroblasts of the ND29369 cell line were subjected to aCGH analysis (Cell Line Genetics, Madison, WI, USA) to determine this cell line’s genotypic variation and the normal karyotype of the iPSCs reprogrammed from this cell line was confirmed in the ND29369 clones 1 and 4 (Cell Line Genetics, Madison, WI, USA) (Supplement) as described previously (Novak et al., 2021).
4.2. Reprogramming
Reprogramming of fibroblasts into pluripotent stem cells was done using Sendai virus at the Yale Human Embryonic Stem Cell Core, New Haven CT, USA. Early passage iPSCs were then passaged and characterized as described previously (Novak et al., 2021).
4.3. iPSC clone classification
The iPSC status was confirmed by immunocytochemistry (Fig. 1a and Supplement), as described previously (Novak et al., 2021). The primary antibodies used were anti-POU5F1 (also known as Oct3/4) and anti-Tra-1-60, and the secondary antibody was Donkey anti-Mouse IgG, Alexa Fluor Plus 555 (Table 3). The PINK1 ILE368ASN mutation (rs774647122, NM_032409.3: 1194 T > A) in the cell line ND40066 (LCSBi002) and the PARKIN R275W mutation (rs34424986, NM_004562.3: 921C > T) in the cell line ND29369 (LCSBi004) were confirmed as described earlier (Novak et al., 2021), using primers listed in Table 3 (Fig. 1c and Supplement). Expression analysis was performed using quantitative PCR (qPCR) as described earlier (Novak et al., 2021) (Fig. 1b) using primers listed in Table 3. The expression is in fg/ul of cDNA sample, where 1 ul of sample represents 0.01ug of total RNA converted to cDNA, as described previously (Novak et al., 2021). The iPSC status and trilineage potential was further confirmed by a TaqMan iPSC Scorecard Assay (ThermoFisher Scientific, Fig. 1d and Supplement) according to the manufacturer’s protocol and as described previously (Novak et al., 2021).
Table 3. Reagents detail.
| Antibodies used for immunocytochemistry | |||
|---|---|---|---|
| Empty Cell | Antibody | Dilution | Company Cat # and RRID |
| Pluripotency Markers | Mouse anti-Human POU5F1 (Oct3/4) | 1:500 | Santa Cruz Biotechnology sc-5279 RRID AB_628051 |
| Mouse anti-Human Tra-1-60 | 1:500 | Merck Millipore MAB4360 RRID AB_2119183 | |
| Secondary antibodies | Donkey anti-Mouse IgG, Alexa Fluor Plus 555 | 1:1000 | ThermoFisher A32773 RRID AB_2762848 |
| Nuclear stain | DAPI | 300 nM | Thermofisher D1306 |
| Primers | Target | Forward/Revers e primer (5′- 3′) | |
| Sequencing primers | PINK1 ILE368ASN | 5′-AACTATCCCTGTACCCTGCG-3′ 5′-CCAAAATCTGCGATCACCAGC-3′ | |
| Sequencing primers | PARKIN R275W | 5′-CCGCCACGTGATTTGCTTAG-3′ 5′-TCTGCAGGACACACTCCTCT-3′ | |
| qPCR primers | OCT4 | 5′-TCGAGAACCGAGTGAGAGG-3′ 5′-GAACCACACTCGGACCACA-3′ | |
| qPCR primers | SOX2 | 5′-GCCGAGTGGAAACTTTTGTCG-3′ 5′-GCAGCGTGTACTTATCCTTCTT-3′ | |
| qPCR primers | NANOG | 5′-TTCCCTCCTCCATGGATCTG-3′ 5′-TGTTTCTTGACTGGGACCTTGTC-3′ | |
| qPCR primers | B2M Beta-2-microglobulin | 5′-GAGTATGCCTGCCGTGTG-3′ 5′-AATCCAAATGCGGCATCT-3′ | |
| Trilineage markers | |||
| Trilineage markers (qPCR) | Ectoderm, Mesoderm, Endoderm | Scorecard – Thermofisher – A15870 | |
5. Additional information
For additional methodological details, including mDA neuron differentiation, and for single cell RNAseq analysis data, please see (Novak et al., 2022).
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
This work was made possible in part by support of S.F. from the Michael J Fox Foundation through Head Start Program and National Institutes of Health (NIH) grants R01 NS124848, RF1 AG058476, R37 NS101996, and P01 AG054407.